
-
线粒体 编辑
线粒体(外文名:mitochondrion)是一种存在于大多数细胞中的由两层膜包被的细胞器,是细胞中制造能量的结构,是细胞进行有氧呼吸的主要场所,被称为"power house"。
线粒体拥有自身的遗传物质和遗传体系,但其基因组大小有限,是一种半自主细胞器。
目录
- 1 基本信息
2 形态特征
3 研究历史
4 主要结构
5 主要功能
6 起源学说
7 遗传学说
8 病理原因
9 衰老介绍
10 疾病介绍
11 相关实验
基本信息
编辑中文名:线粒体
分布:大多数真核细胞
分布区域:分布在细胞功能旺盛的区域
作用:为细胞的各种生命活动提供能量
外文名:Mitochondrion
直径:0.5到1.0微米左右
形状:短棒状、圆球状
形态特征
编辑 线粒体结构示意图
线粒体结构示意图
形状
 图中编号为9的是线粒体
图中编号为9的是线粒体
数量
不同生物的不同组织中线粒体数量的差异是巨大的。有许多细胞拥有多达数千个的线粒体(如肝脏细胞中有1000-2000个线粒体),而一些细胞则只有一个线粒体(如酵母菌细胞的大型分支线粒体)。大多数哺乳动物的成熟红细胞不具有线粒体。一般来说,细胞中线粒体数量取决于该细胞的代谢水平,代谢活动越旺盛的细胞线粒体越多。
分布
线粒体分布方向与微管一致,通常分布在细胞功能旺盛的区域:如在肾脏细胞中靠近微血管,呈平行或栅状排列;在肠表皮细胞中呈两极分布,集中在顶端和基部;在精子中分布在鞭毛中区。在卵母细胞体外培养中,随着细胞逐渐成熟,线粒体会由在细胞周边分布发展成均匀分布。线粒体在细胞质中能以微管为导轨、由马达蛋白提供动力向功能旺盛的区域迁移。
组成
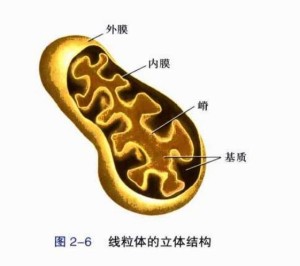
结构
线粒体由外至内可划分为线粒体外膜(OMM)、线粒体膜间隙、线粒体内膜(IMM)和线粒体基质四个功能区。处于线粒体外侧的膜彼此平行,都是典型的单位膜。其中,线粒体外膜较光滑,起细胞器界膜的作用;线粒体内膜则向内皱褶形成线粒体嵴,负担更多的生化反应。这两层膜将线粒体分出两个区室,位于两层线粒体膜之间的是线粒体膜间隙,被线粒体内膜包裹的是线粒体基质。
研究历史
编辑
1857年,瑞士解剖学家及生理学家阿尔伯特·冯·科立克在肌肉细胞中发现了颗粒状结构。另外的一些科学家在其他细胞中也发现了同样的结构,证实了科立克的发现。德国病理学家及组织学家理查德·阿尔特曼将这些颗粒命名为“原生粒”(bioblast)并于1886年发明了一种鉴别这些颗粒的染色法。阿尔特曼猜测这些颗粒可能是共生于细胞内的独立生活的细菌。
1898年,德国科学家卡尔·本达因这些结构时而呈线状时而呈颗粒状,所以用希腊语中“线”和“颗粒”对应的两个词——“mitos”和“chondros”——组成“mitochondrion”来为这种结构命名,这个名称被沿用至今。一年后,美国化学家莱昂诺尔·米歇利斯开发出用具有还原性的健那绿染液为线粒体染色的方法,并推断线粒体参与某些氧化反应。这一方法于1900年公布,并由美国细胞学家埃德蒙·文森特·考德里推广。德国生物化学家奥托·海因里希·沃伯格成功完成线粒体的粗提取且分离得到一些催化与氧有关的反应的呼吸酶,并提出这些酶能被氰化物(如氢氰酸)抑制的猜想。
英国生物学家大卫·基林在1923年至1933年这十年间对线粒体内的氧化还原链(redox chain)的物质基础进行探索,辨别出反应中的电子载体——细胞色素。
沃伯格于1931年因“发现呼吸酶的性质及作用方式”被授予诺贝尔生理学或医学奖。
美国弗吉尼亚大学最新一项研究表明,动植物细胞中的线粒体其实是寄生细菌,早期寄生细菌可以对动物和植物提供能量,在细胞中作为能量寄生虫存在,对寄居体十分有益。新一代DNA序列技术解码18种细菌基因组,这些细菌是线粒体的近亲生物。
主要结构
编辑 线粒体剖面图
线粒体剖面图
膜间隙
线粒体膜间隙是线粒体外膜与线粒体内膜之间的空隙,宽约6-8nm,其中充满无定形液体。由于线粒体外膜含有孔蛋白,通透性较高,而线粒体内膜通透性较低,所以线粒体膜间隙内容物的组成与细胞质基质十分接近,含有众多生化反应底物、可溶性的酶和辅助因子等。线粒体膜间隙中还含有比细胞质基质中浓度更高的腺苷酸激酶、单磷酸激酶和二磷酸激酶等激酶,其中腺苷酸激酶是线粒体膜间隙的标志酶。线粒体膜间隙中存在的蛋白质可统称为“线粒体膜间隙蛋白质”,这些蛋白质全部在细胞质基质中合成。
内膜
线粒体内膜是位于线粒体外膜内侧、包裹着线粒体基质的单位膜。线粒体内膜中蛋白质与磷脂的质量比约为0.7:0.3,并含有大量的心磷脂(心磷脂常为细菌细胞膜的成分)。线粒体内膜的某些部分会向线粒体基质折叠形成线粒体嵴。线粒体内膜的标志酶是细胞色素氧化酶。
线粒体通过向内凹形成嵴,从而来增加内膜面积。然后是更多的反应能在内膜上进行。
线粒体内膜含有比外膜更多的蛋白质(超过151种,约占线粒体所含所有蛋白质的五分之一),所以承担着更复杂的生化反应。存在于线粒体内膜中的几类蛋白质主要负责以下生理过程:特异性载体运输磷酸、谷氨酸、鸟氨酸、各种离子及核苷酸等代谢产物和中间产物;内膜转运酶(translocase of the inner membrane,TIM)运输蛋白质;参与氧化磷酸化中的氧化还原反应;参与ATP的合成;控制线粒体的分裂与融合。
嵴
线粒体嵴简称“嵴”,是线粒体内膜向线粒体基质折褶形成的一种结构。线粒体嵴的形成增大了线粒体内膜的表面积。在不同种类的细胞中,线粒体嵴的数目、形态和排列方式可能有较大差别。线粒体嵴主要有几种排列方式,分别称为“片状嵴”(lamellar cristae)、“管状嵴”(tubular cristae)和“泡状嵴”(vesicular cristae)。片状排列的线粒体嵴主要出现在高等动物细胞的线粒体中,这些片状嵴多数垂直于线粒体长轴;管状排列的线粒体嵴则主要出现在原生动物和植物细胞的线粒体中。有研究发现,睾丸间质细胞中既存在层状嵴也存在管状嵴。线粒体嵴上有许多有柄小球体,即线粒体基粒,基粒中含有ATP合酶,能利用呼吸链产生的能量合成三磷酸腺苷。所以需要较多能量的细胞,线粒体嵴的数目一般也较多。但某些形态特殊的线粒体嵴由于没有ATP合酶,所以不能合成ATP。
基质
 膜结构
膜结构
线粒体DNA是线粒体中的遗传物质,呈双链环状。一个线粒体中可有一个或数个线粒体DNA分子。线粒体RNA是线粒体DNA的表达产物,RNA编辑也普遍存在于线粒体RNA中,是线粒体产生功能蛋白所必不可少的过程。线粒体核糖体是存在于线粒体基质内的一种核糖体,负责完成线粒体内进行的翻译工作。线粒体核糖体的沉降系数介干55S-56S之间。一般的线粒体核糖体由28S核糖体亚基(小亚基)和39S核糖体亚基(大亚基)组成。在这类核糖体中,rRNA约占25%,核糖体蛋白质约占75%。线粒体核糖体是已发现的蛋白质含量最高的一类核糖体。线粒体基质中存在的蛋白质统称为“线粒体基质蛋白质”,包括DNA聚合酶、RNA聚合酶、柠檬酸合成酶以及三羧酸循环酶系中的酶类。大部分线粒体基质蛋白是由核基因编码的。线粒体基质蛋白不一定只在线粒体基质中表达,它们也可以在线粒体外表达。
主要功能
编辑线粒体是真核生物进行氧化代谢的部位,是糖类、脂肪和氨基酸最终氧化释放能量的场所。线粒体负责的最终氧化的共同途径是三羧酸循环与氧化磷酸化,分别对应有氧呼吸的第二、三阶段。细胞质基质中完成的糖酵解和在线粒体基质中完成的三羧酸循环在会产还原型烟酰胺腺嘌呤二核苷酸(reduced nicotinarnide adenine dinucleotide,NADH)和还原型黄素腺嘌呤二核苷酸(reduced flavin adenosine dinucleotide,FADH2)等高能分子,而氧化磷酸化这一步骤的作用则是利用这些物质还原氧气释放能量合成ATP。在有氧呼吸过程中,1分子葡萄糖经过糖酵解、三羧酸循环和氧化磷酸化将能量释放后,可产生30-32分子ATP(考虑到将NADH运入线粒体可能需消耗2分子ATP)。如果细胞所在环境缺氧,则会转而进行无氧呼吸。此时,糖酵解产生的丙酮酸便不再进入线粒体内的三羧酸循环,而是继续在细胞质基质中反应(被NADH还原成乙醇或乳酸等发酵产物),但不产生ATP。所以在无氧呼吸过程中,1分子葡萄糖只能在第一阶段产生2分子ATP。
三羧酸循环
糖酵解中生成的每分子丙酮酸会被主动运输转运穿过线粒体膜。进入线粒体基质后,丙酮酸会被氧化,并与辅酶A结合生成CO2、还原型辅酶Ⅰ和乙酰辅酶A。乙酰辅酶A是三羧酸循环(也称为“柠檬酸循环”或“Krebs循环”)的初级底物。参与该循环的酶除位于线粒体内膜的琥珀酸脱氢酶外都游离于线粒体基质中。在三羧酸循环中,每分子乙酰辅酶A被氧化的同时会产生起始电子传递链的还原型辅因子(包括3分子NADH和1分子FADH2)以及1分子三磷酸鸟苷(GTP)。
氧化磷酸化
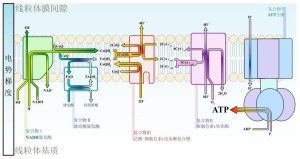
当质子被泵入线粒体膜间隙后,线粒体内膜两侧便建立起了电化学梯度,质子就会有顺浓度梯度扩散的趋势。质子唯一的扩散通道是ATP合酶(呼吸链复合物V)。当质子通过复合物从膜间隙回到线粒体基质时,电势能被ATP合酶用于将ADP和磷酸合成ATP。这个过程被称为“化学渗透”,是一种协助扩散。彼得·米切尔就因为提出了这一假说而获得了1978年诺贝尔奖。1997年诺贝尔奖获得者保罗·博耶和约翰·瓦克阐明了ATP合酶的机制。
储存钙离子
线粒体可以储存钙离子,可以和内质网、细胞外基质等结构协同作用,从而控制细胞中的钙离子浓度的动态平衡。线粒体迅速吸收钙离子的能力使其成为细胞中钙离子的缓冲区。在线粒体内膜膜电位的驱动下,钙离子可由存在于线粒体内膜中的单向运送体输送进入线粒体基质;排出线粒体基质时则需要钠-钙交换蛋白的辅助或通过钙诱导钙释放(calcium-induced-calcium-release,CICR)机制。在钙离子释放时会引起伴随着较大膜电位变化的“钙波”(calcium wave),能激活某些第二信使系统蛋白,协调诸如突触中神经递质的释放及内分泌细胞中激素的分泌。线粒体也参与细胞凋亡时的钙离子信号转导。
其他功能
除了合成ATP为细胞提供能量等主要功能外,线粒体还承担了许多其他生理功能。
 显微镜下
显微镜下
·细胞增殖与细胞代谢的调控;
·合成胆固醇及某些血红素。
线粒体的某些功能只有在特定的组织细胞中才能展现。例如,只有肝脏细胞中的线粒体才具有对氨气(蛋白质代谢过程中产生的废物)造成的毒害解毒的功能。
起源学说
编辑 线粒体
线粒体
内共生学说
该学说认为线粒体起源于被另一个细胞吞噬的线粒体祖先——原线粒体——一种能进行三羧酸循环和电子传递的革兰氏阴性菌。这种好氧细菌是变形菌门下的一个分支,与立克次氏体有密切关系。原线粒体被吞噬后,并没有被消化,而是与宿主细胞形成了共生关系——寄主可以从宿主处获得更多营养,而宿主则可使用寄主产生的能量——这种关系增加了细胞的竞争力,使其可以适应更多的生存环境。在长期对寄主和宿主都有利的互利共生中,原线粒体逐渐演变形成了线粒体,使宿主细胞中进行的糖酵解和原线粒体中进行的三羧酸循环和氧化磷酸化成功耦合。有研究认为,这种共生关系大约发生在17亿年以前,与进化趋异产生真核生物和古细菌的时期几乎相同。但线粒体与真核生物细胞核出现的先后关系仍存在争议。
现已发现支持内共生学说的证据包括:
- 遗传信息转移:近期的分子生物学和生物信息学的研究发现真核细胞的细胞核中存在可能属于呼吸细菌或蓝细菌的遗传信息,说明最初的呼吸细菌和蓝细菌的大部分基因组在漫长的共进化过程中发生了向细胞核的转移。
- 线粒体基因组与细菌基因组具有明显的相似性:包括1)线粒体拥有自己DNA,其形状与细菌的环状DNA类似,无组蛋白结合;2)碱基比例、核苷酸序列、基因结构特征等方面相似,不含5mC;3)线粒体具有自身的DNA聚合酶及RNA聚合酶,能进行独立的复制与转录;4)其mRNA、rRNA的沉降系数与细菌的相似。
- 线粒体具备独立、完整的蛋白质合成系统:与真核细胞的蛋白质合成系统相比,线粒体蛋白质合成的多数特征与细菌蛋白质合成系统更相似,包括1)蛋白质合成从N-甲酰甲硫氨酸开始,而真核细胞从甲硫氨酸开始;2)线粒体的核糖体小于真核生物的80S核糖体;3)线粒体、叶绿体、原核生物中存在5SrRNA,而不少真核生物的核糖体中存在5.8SrRNA;4)线粒体中的蛋白质合成因子具有原核生物核糖体的识别特异性,但不能识别细胞质核糖体;5)线粒体mRNA与线粒体核糖体形成多核糖体;6)线粒体、叶绿体上的蛋白质合成可被氯霉素、四环素所抑制,而抑制真核生物蛋白质合成的放线菌酮则对他们无抑制作用;7)线粒体的RNA聚合酶可被原核细胞RNA聚合酶抑制剂利福霉素所抑制,但不被真核细胞RNA聚合酶抑制剂放线菌素D所抑制等。
- 线粒体分裂方式与细菌相似:线粒体及叶绿体均以缢裂的方式分裂增殖,类似于细菌。
- 线粒体的膜特性:线粒体外膜与真核细胞内膜相似,线粒体内膜与细菌质膜相似;线粒体内膜的蛋白质/脂质的比例远大于外膜,与细菌相似。
- 其他特征:线粒体的磷脂成分、呼吸类型和Cyt c的初级结构均与反硝化副球菌或紫色非硫光合细菌非常接近,暗示线粒体的祖先可能是这两种菌的一种。
- 遗传密码比较:线粒体的遗传密码与变形菌门细菌的遗传密码更为相似;
不足之处:
从进化角度,如何解释在代谢上明显占优势的共生体反而将大量的遗传信息转移到宿主细胞中?
不能解释细胞核是如何进化来的,即原核细胞如何演化为真核细胞?
线粒体和叶绿体的基因组中存在内含子,而真细菌原核生物基因组中不存在内含子,如果同意内共生起源学说的观点,那么线粒体和叶绿体基因组中的内含子从何发生?
非内共生学说
非内共生学说又称为“细胞分化学说”,认为线粒体的发生是由细胞膜或内质网膜等生物膜系统中的膜结构演变而来的。非内共生学说有几种模型,主流的模型认为在细胞进化的最初阶段,原核细胞基因组复制后并不伴有典型的无丝分裂,而是拟核附近的细胞膜内陷形成双层膜,将其中一个基因组包围、隔离,进而发生细胞分裂。未分裂出来的子细胞则缓慢演化为细胞核、线粒体和叶绿体等高度特化的细胞结构。
不足之处:
实验证据不多
无法解释为何线粒体、叶绿体与细菌在DNA分子结构和蛋白质合成性能上有那么多相似之处
对线粒体和叶绿体的DNA酶、RNA酶和核糖体的来源也很难解释。
真核细胞的细胞核能否起源于细菌的核区?
遗传学说
编辑 线粒体
线粒体
| 生物 | 学名 | mtDNA长度(bp) |
|---|---|---|
| 芽殖酵母 | Saccharomyces cerevisiae | 85779 |
| 裂殖酵母 | Schizosaccharomyces pombe | 19431 |
| 拟南芥 | Arabidopsis thaliana | 366924 |
| 水稻 | Oryza sativa | 490520 |
| 秀丽隐杆线虫 | Caenorhabditis elegans | 13794 |
| 黑腹果蝇 | Drosophila melanogaster | 19517 |
| 非洲爪蟾 | Xenopus laevis | 17553 |
| 小鼠 | Mus musculus | 16300 |
mtDNA利用率极高,线粒体基因组各基因之间排列十分紧凑,部分区域还可能出现重叠(即前一个基因的最后一段碱基与下一个基因的第一段碱基相衔接)。人类mtDNA中基因间隔区总共只有87bp,占mtDNA总长的0.5%。mtDNA的两条DNA单链均有编码功能,其中重链编码两个rRNA、12个mRNA和14个tRNA;轻链编码一个mRNA和8个tRNA。mtDNA一般没有内含子(如人类的mtDNA等),但也已发现某些真核生物的mtDNA拥有内含子,这些生物包括:盘基网柄菌等原生生物和酵母菌(其OXi3基因有9个内含子)。这些mtDNA中的内含子在基因转录产物的加工和翻译中可能有一定功能。
线粒体基因组通常都是存在于同一个mtDNA分子中,但少数生物的线粒体基因组却分别储存在多个不同的mtDNA中。例如,人虱的线粒体基因组就分开储藏于18个长约3-4kb的微型环状DNA中,每个DNA分子只分配到了1-3个基因。这些微型环状DNA之间也存在着同源或非同源的基因重组现象,但成因未知。
遗传密码
线粒体中拥有一套独特的遗传系统。在进行人类线粒体遗传学研究时,人们确认线粒体的遗传密码与通用遗传密码也有些许差异。自从上述发现证明并不只存在单独的一种遗传密码之后,许多有轻微不同的遗传密码都陆续连发现。在线粒体的遗传密码中最常见的差异是:AUA由终止密码子变为甲硫氨酸的密码子、UGA由终止密码子变为色氨酸的密码子、AGA和AGG由精氨酸的密码子变为终止密码子(植物等生物的线粒体遗传密码另有差异,参见表二)。此外,也有某些特例是只涉及终止密码子的,在山羊支原体线粒体遗传密码的UGA由终止密码子变为色氨酸的密码子,而且使用频率比UGG更高;四膜虫线粒体遗传密码里只有UGA一种终止密码子,其UAA和UAG由终止密码子变为谷氨酰胺的密码子;而游仆虫线粒体遗传密码里则只有UAA和UAG两种终止密码子,其UGA由终止密码子变为半胱氨酸的密码子。通过线粒体遗传密码和通用遗传密码的对比,可以推导出遗传密码演化过程的可能模式。
| 密码子 | 通用密码 | 线粒体遗传密码 | |||
| 真菌 | 植物 | 无脊椎动物 | 哺乳动物 | ||
| UGA | 终止密码子 | 色氨酸 | 终止密码子 | 色氨酸 | 色氨酸 |
| AUA | 异亮氨酸 | 甲硫氨酸 | 异亮氨酸 | 甲硫氨酸 | 甲硫氨酸 |
| CUA | 亮氨酸 | 苏氨酸 | 亮氨酸 | 亮氨酸 | 亮氨酸 |
| AGA、AGG | 精氨酸 | 精氨酸 | 精氨酸 | 丝氨酸 | 终止密码子 |
分裂与融合
线粒体的融合是与分裂协同进行的,过程高度保守,需要在多种蛋白质的精确调控下完成。两者一般保持动态平衡,这种平衡对维持线粒体正常的形态、分布和功能十分重要。线粒体融合与分裂间的失衡可产生巨型线粒体,这种过大的线粒体常见于病变的肝细胞、恶性营养不良患者的胰脏细胞和白血病患者骨髓的巨噬细胞中。分裂异常会导致线粒体破碎,而融合异常则会导致线粒体形态延长,两者都会影响线粒体的功能。分裂与融合活动异常的线粒体膜电位通常会降低,并最终经线粒体自噬作用清除。
线粒体的分裂在真核细胞内经常发生。为了保证在细胞发生分裂后每个子细胞都能继承母细胞的线粒体,母细胞中的线粒体在一个细胞周期需要至少复制一次。即使是在不再分裂的细胞内,线粒体为了填补已老化的线粒体造成的空缺也需要进行分裂。的线粒体以与细菌的无丝分裂类似的方式进行增殖,可细分为三种模式:
间壁分离(见于部分动物和植物线粒体):线粒体内部首先由内膜形成隔,随后外膜的一部分内陷,插入到隔的双层膜之间,将线粒体一分为二。
收缩分离(见于蕨类植物和酵母菌线粒体):线粒体中部先缢缩同时向两端不断拉长然后一分为二。
出芽分离(见于藓类植物和酵母菌线粒体):线粒体上先出现小芽,小芽脱落后成长、发育为成熟线粒体。
线粒体的融合也是细胞中的基本事件,对线粒体正常功能的发挥具有非常重要的作用。人类细胞需要通过线粒体融合的互补作用来抵抗衰老;酵母细胞线粒体融合发生障碍会引起呼吸链缺陷。线粒体间的融合需在一种分子量约为800kDa的蛋白质复合物——“融合装置”(fusion machinery)的介导下进行,该过程可大致分为四个步骤:锚定、外膜融合、内膜融合以及基质内含物融合。
群体遗传学
因为mtDNA几乎不发生基因重组,所以遗传学家长期将其作为研究群体遗传学与进化生物学的信息来源。所有mtDNA是以单一单元(单体型)进行遗传的(而不像细胞核中的DNA储存在多个染色体中),它们在亲本与子代之间的传递关系并不复杂,因此不同个体间mtDNA的联系便可以利用系统发生树来表现。而从这些系统发生树的形态中人们可以得知种群的进化史。人类进化遗传学中运用分子钟技术推算出了线粒体夏娃最晚出现的时间(这个成果被认为是人类由非洲单地起源的有力依据)是利用mtDNA研究群体遗传学的典型例子。另外一个例子是对尼安德特人骨骼化石中mtDNA测序。该测序的结果显示,尼安德特人与解剖学意义上的现代人在mtDNA序列上有较大差异,说明两者间缺乏基因交流。虽然mtDNA在遗传学研究中占据了重要地位,但是mtDNA序列中的信息只能反映所考察的群体中的雌性成员的演化进程,而不能代表整个种群。这一缺陷需要由对父系遗传序列(如Y染色体上的非重组区)的测序弥补。广义上来说,只有既考虑了mtDNA又考虑了核DNA的遗传学研究才能为种群的进化史提供全面的线索。
病理原因
编辑 线粒体
线粒体
数量的改变
线粒体的平均寿命约为10天。衰亡的线粒体可通过保留的线粒体直接分裂为二予以补充。在病理状态下,线粒体的增生实际上是对慢性非特异性细胞损伤的适应性反应或细胞功能升高的表现。例如心瓣膜病时的心肌线粒体、周围血液循环障碍伴间歇性跛行时的骨骼肌线粒体的呈增生现象。
线粒体数量减少则见于急性细胞损伤时线粒体崩解或自溶的情况下,持续约15分钟。慢性损伤时由于线粒体逐渐增生,故一般不见线粒体减少(甚至反而增多)。此外,线粒体的减少也是细胞未成熟和(或)去分化的表现。
大小改变
细胞损伤时最常见的改变为线粒体肿大。根据线粒体的受累部位可分为基质型肿胀和嵴型肿胀二种类型,而以前者为常见。基质型肿胀时线粒体变大变圆,基质变浅、嵴变短变少甚至消失(图1-9)。在极度肿胀时,线粒体可转化为小空泡状结构。此型肿胀为细胞水肿的部分改变。光学显微镜下所谓的浊肿细胞中所见的细颗粒即肿大的线粒体。嵴型肿较少见,此时的肿胀局限于嵴内隙,使扁平的嵴变成烧瓶状乃至空泡状,而基质则更显得致密。嵴型肿胀一般为可复性,但当膜的损伤加重时,可经过混合型而过渡为基质型。
线粒体为对损伤极为敏感的细胞器,其肿胀可由多种损伤因子引起,其中最常见的为缺氧;此外,微生物毒素、各种毒物、射线以及渗透压改变等亦可引起。但轻度肿大有时可能为其功能升高的表现,较明显的肿胀则恒为细胞受损的表现。但只要损伤不过重、损伤因子的作用不过长,肿胀仍可恢复。
线粒体的增大有时是器官功能负荷增加引起的适应性肥大,此时线粒体的数量也常增多,例如见于器官肥大时。反之,器官萎缩时,线粒体则缩小、变少。
结构的改变
线粒体嵴是能量代谢的明显指征,但嵴的增多未必均伴有呼吸链酶的增加。嵴的膜和酶平行增多反映细胞的功能负荷加重,为一种适应状态的表现;反之,如嵴的膜和酶的增多不相平行,则是胞浆适应功能障碍的表现,此时细胞功能并不升高。
在急性细胞损伤时(大多为中毒或缺氧),线粒体的嵴被破坏;慢性亚致死性细胞损伤或营养缺乏时,线粒体的蛋白合成受障,以致线粒体几乎不再能形成新的嵴。
根据细胞损伤的种类和性质,可在线粒体基质或嵴内形成病理性包含物。这些包含物有的呈晶形或副晶形(可能由蛋白构成),如在线粒体性肌病或进行性肌营养不良时所见,有的呈无定形的电子致密物,常见于细胞趋于坏死时,乃线粒体成分崩解的产物(脂质和蛋白质),被视为线粒体不可复性损伤的表现。线粒体损伤的另一种常见改变为髓鞘样层状结构的形成,这是线粒体膜损伤的结果。
衰亡或受损的线粒体,最终由细胞的自噬过程加以处理并最后被溶酶体酶所降解消化。
衰老介绍
编辑线粒体是直接利用氧气制造能量的部位,90%以上吸入体内的氧气被线粒体消耗掉。但是,氧是个“双刃剑”,一方面生物体利用氧分子制造能量,另一方面氧分子在被利用的过程中会产生极活泼的中间体(活性氧自由基)伤害生物体造成氧毒性。生物体就是在不断地与氧毒性进行斗争中求得生存和发展的,氧毒性的存在是生物体衰老的最原初的原因。线粒体利用氧分子的同时也不断受到氧毒性的伤害,线粒体损伤超过一定限度,细胞就会衰老死亡。生物体总是不断有新的细胞取代衰老的细胞以维持生命的延续,这就是细胞的新陈代谢。
疾病介绍
编辑人类线粒体出现问题会导致线粒体病,线粒体病是一大类遗传代谢病,线粒体病主要包括:母系遗传Leigh综合征、线粒体肌病、多系统疾病、心肌病、进行性眼外肌麻痹、Leer遗传性视神经病、线粒体肌病,肌病,糖尿病和耳聋、共济失调舞蹈病、细胞外基质慢性游走性红斑、进行性眼外肌麻痹、肌红蛋白尿电机神经元疾病,铁粒幼细胞贫血、MERRF-线粒体肌病、肌阵挛(癫痫)、线粒体脑肌病、MERRF、线粒体肌病、共济失调并发色素性视网膜炎、家族性双侧纹状体坏死、共济失调并发色素性视网膜炎、家族性双侧纹状体坏死、骨骼肌溶解症、婴儿猝死综合征等等疾病。
线粒体病遗传方式复杂,导致疾病的原因主要由核基因和线粒体基因造成,临床表现复杂,确切病因的诊断十分困难,往往通过大分子酶学活性检测分析并结合遗传学基因分析的双重手段确定病因。
线粒体基因组属于母系遗传,为了避免新生儿缺陷,产前妈妈的线粒体基因组分析十分必要。
2014年7月,科学家发现促进癌症转移的线粒体开关,线粒体是细胞的能量工厂,当肿瘤细胞中线粒体的功能发生改变时就会促进细胞的迁移,最终导致肿瘤成功转移。研究人员测定了肿瘤细胞中线粒体促进肿瘤转移过程中涉及的分子机制,结果发现,在特定的条件下,线粒体可以产生过多的超氧离子自由基,超氧离子的过量产生就会引发肿瘤转移灶的形成,最终肿瘤转移组织就会在新的组织中形成肿瘤。
相关实验
编辑线粒体——示教:3号片
小狗胰脏,Regaud氏液固定,石蜡切片,铁苏木素染色。
线粒体用铁苏木素染色呈黑色,分布于核周围的细胞质中,线粒体在高倍镜下呈粒状、线状或短棒状,或直或曲,轮廓鲜明。
胰脏的分泌细胞呈锥形,核大而圆,位于细胞中央,细胞游离端聚集有许多大而圆的黑色颗粒为分泌颗粒。
提取观察
线粒体是细胞中重要的细胞器,存在于绝大多数生活细胞中,它的主要功能是提供细胞内各种物质代谢所需要的能量。正由于这样,对线粒体膜,呼吸链酶及线粒体DNA等成分的结构,功能以及物理化学性质的研究已经成为细胞生物学研究中的重要课题,所以提取线粒体的技术已经成为线粒体研究中必不可少的手段,线粒体大量存在于代谢旺盛的细胞中,如动物的心肌,肝,肾等器官和组织的细胞中,大量置备线粒体就是从这些器官组织中提取,当所用样品较少时(如电镜和光镜的观察)可采用从组织培养细胞中提取,本实验就是介绍两种材料制备用于光镜观察的线粒体。
一、目的与要求
了解提取线粒体的基本原理及其过程,通过光学显微镜的观察了解体外分离的线粒体的一般形态。
二、基本原理
线粒体具有完整的结构,一定的大小和质量,低温条件下在等渗液中破碎细胞,差速离心后,获得线粒体。经活性染料健那绿Janus green B染色,线粒体呈浅蓝色。
三、实验内容
1.线粒体的分离提取 2. 鼠肝的匀浆制备 3. 线粒体的活体染色
四、实验步骤
(一)动物组织线粒体的分离,提取与观察
显微镜检查:将1%Janus green B溶液按1:1比例加入线粒体悬液中,在室温或水浴中染15~20分钟,用吸管吸取一滴线粒体悬液,滴于载玻片上,加盖玻片后,放显微镜下进行观察,线粒体为蓝绿色圆形颗粒。
2.组织培养细胞的线粒体的提取与观察
注意问题
1.整个操作过程为保证线粒体的完整,应尽量使操作时的环境如温度(0—4℃),pH (7.0左右)保持恒定,同时尽可能短操作时间。
2.组培细胞消化时要特别小心,防止损失或反复。(损失指细胞脱落到消化液中)。
3.匀浆时,所用的介质一定是等渗缓冲液,常用的有0.25 mol/L蔗糖溶液或生理盐水代替Hank’s液
4.匀浆次数依照匀浆器的松紧而定,次数过少,细胞破损不完全,就会影响线粒体产量。
5.所以取2/3上清夜用来制备线粒体是为防止细胞碎片过多影响观察。
6.整个分离过程,一般最好在30—60分钟内完成,不宜过长。















-

北条麻妃
北条麻妃,原名白石さゆり,以其高挑的身材以及贵族的气质称霸熟女界,1974年3月26日出生于日本石川县,AV女优。
2024-08-18 -

橘ひなた
橘日向(橘 ひなた)1990年8月12日出生,是日本90后AV女优。 2009年11月出道,已经下马拍了多部步兵。
2023-01-10 -

武藤兰
武藤兰,1980年9月4日b出生,初期曾使用过清水优香的艺名,后名武藤兰,日本AV女优。
2023-01-07 -

成濑心美
成濑心美,1989年3月3日出生于日本新潟县,E罩杯,2008年12月以素人身份出道,日本AV女优。
2023-01-07 -

波多野结衣
波多野结衣(はたの ゆい),女,1988年5月24日出生于日本京都府,著名日本女演员、AV女优。
2023-01-07
